![]()
19 février 2013 - Journée d'étude ASFC «Sharm
El-Sheikh - Ombres & Lumières»
Nutrition
et Physiologie digestive
Les apports lors du 10ème Congrès Mondial de Cuniculture
par
Thierry GIDENNE * et Jean-Marc SALAÜN **
* INRA Toulouse, UMR 1289 Tandem, CS 52627, 31326 Castanet-Tolosan
** Sanders, Centre d'affaire Odyssée - Cicé Blossac - CS17228,
35172 Bruz.

J.M. Salaün lors de la présentation du rapport
Comme les années passées, la section "Alimentation, nutrition et physiologie de la digestion" a été celle contenant le plus de communications courtes. Elle représente à elle seule 22% des travaux présentés lors de ce 10ème Congrès, soit 42 communications (sur un total de 194) et 2 synthèses (sur un total de 8 synthèses).
Dans cet article,
nous traiterons des résultats nouveaux en nutrition (nutriments
et recommandations), en physiologie de la digestion, et des effets de
divers additifs alimentaires sur la physiologie, ce qui correspond à
l'analyse de 19 communications courtes et des 2 synthèses. Les
résultats sur les techniques d'alimentation (y compris le rationnement)
et la valeur nutritive des matières premières sont traités
dans l'article de MM Lebas et Duperray.
|
Trois études ont utilisé des outils de microbiologie moléculaire afin d'analyser le microbiote caecal (Vantus et al.; Kimsé et al.; El Abed et al.). Après extraction et amplification (PCR) de l'ADN des bactéries il est possible de comparer les différentes séquences d'ADN, et de mesurer "l'empreinte" ou fingerprint du microbiote. A partir du profil bactérien, il est ensuite possible d'estimer un index de la biodiversité (combinaison de la richesse et de l'abondance), et aussi d'analyser la structure de la communauté microbienne (c'est à dire déterminer si les espèces présentes sont les mêmes dans 2 échantillons). Rappelons que l'un des avantages de la microbiologie moléculaire est de prendre en compte la quasi-totalité des microorganismes présents, et pas seulement les 20 à 30% cultivables. La comparaison des séquences d'ADN obtenues avec celles de banques de données permet également d'identifier la famille voire le genre des bactéries. Ainsi, dans la synthèse de Combes et al., on constate que les progrès dans les techniques de séquençage permettent de préciser l'affiliation taxonomique des bactéries, dans un phylum (pour au moins 95% des séquences), dans une famille (pour au moins 50% des séquences) ou dans un genre (10 à 30 % des séquences) |
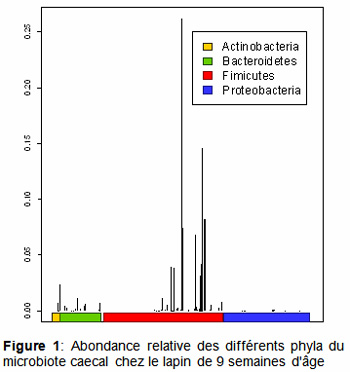 |
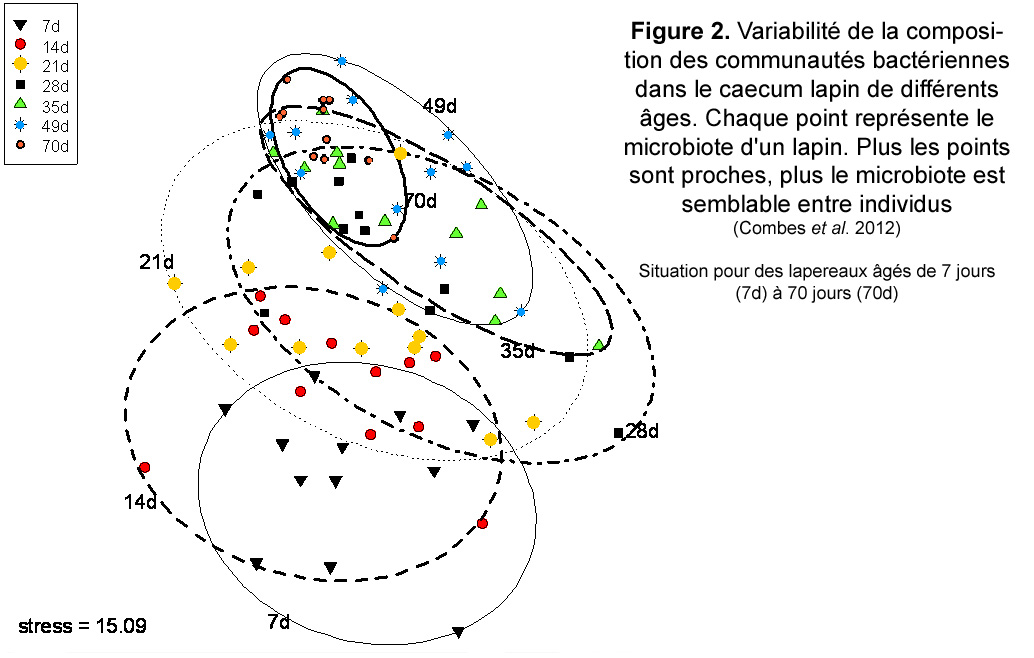
Figure 2. Variabilité de la composition des communautés bactériennes dans le caecum lapin de différents âges. Chaque point représente le microbiote d'un lapin. Plus les points sont proches, plus le microbiote est semblable entre individus
Le phylum des Firmicutes est très majoritaire (abondance relative = 90%) dans le microbiote caecal du lapin en fin de croissance (figure 1, Combes et al.) , suivi par le phylum Bacteroidetes (abondance < 5%).
Cependant, entre deux
semaines et l'âge adulte, le microbiote caecal évolue très
sensiblement, avec une assez grande diversité de profils bactériens
entre les individus jeunes, puis une homogénéisation des
profils avec l'âge (figure 2, Combes et al.). Ainsi, Vantus
et al., montrent par qPCR, que l'abondance relative de Bacteroidetes et
de 2 types de Clostridium (leptum et cocoides, appartenant au phylum des
Firmicutes) n'évolueraient plus entre 49 et 80 jours d'âge.
Il serait donc possible d'intervenir pour modifier l'implantation du microbiote
caecal du lapin, dans une fenêtre de temps entre une et quatre semaines
d'âge, là où la variabilité de microbiote entre
individus est encore élevée. Au delà, on peut penser
que le microbiote qui est implanté est de plus en plus semblable
entre individus, et donc plus stable et difficile à modifier.
Ainsi, Combes et al. ont décrit plusieurs voies d'action pour orienter le microbiote vers un service plus favorable à l'hôte, comme : agir très précocement sur le lapereau encore au nid (moduler l'environnement immédiat dans le nid, etc...), ou agir sur la nutrition avant sevrage ou juste après sevrage (les fibres devraient jouer un rôle important, sans exclure les autres nutriments, ou l'usage de probiotiques).
.Des prébiotiques, tels que des Mannanoligosaccharides (MOS) ou des ?-glucanes semblerait globalement modifier le microbiote caecal et iléal, étudié par une technique d'empreinte moléculaire (T-RFLP; El Abed et al.). Avec une méthode proche (SSCP), Kimsé et al. constatent que l'addition d'un probiotique (levures) accroît légèrement la biodiversité du microbiote caecal.
L'addition de MOS stimulerait l'activité microbienne caecale selon Iannaccone et al., tandis que l'addition d'un cocktail d'antibiotiques (colistine+tylosine+oyxytetracycline) l'inhiberait. Par rapport au témoin ou au groupe recevant des antibiotiques, l'addition de MOS conduirait également à une plus faible mortalité post-sevrage (21 et 20% vs 14% respectivement)
Par ailleurs, on peut signaler que l'addition de spiruline (micro-algue) ou de thym n'affecte pas l'activité microbienne caecale (ni le développement digestif) du lapin en croissance ni l'abondance des bactéries totales ou anaérobie stricte (Bonai et al.).
Enfin, une équipe
Tchèque (Marounek et al.) a comparé l'activité
fermentaire caecale chez le lièvre (sauvage) et le lapin domestique.
Ils constatent une plus forte concentration en AGV activité chez
le lièvre (mais plus d'NH3), probablement en lien avec son régime;
mais aussi le profil fermentaire chez le lièvre présente
un ratio C3/C4 supérieur à 1, contrairement au lapin. En
culture in vitro, il semble que la production de méthane
soit significative chez le lapin, mais presque nulle chez le lièvre.
Par une technique
de cannulation du canal pancréatique chez le lapin adulte anesthésié,
Dojana et al. ont mesuré d'une part le flux global de suc
pancréatique et aussi l'activité de certaines enzymes, en
fonction du régime. Ainsi, la sécrétion pancréatique
globale n'est pas modifiée par l'alimentation; en revanche ils
constatent assez logiquement, une plus forte activité de l'amylase
avec un régime "amylacé", de même pour la
trypsine avec le régime riche en protéines, alors que l'ajout
de lipides n'a pas d'effet significatif sur la lipase ou les autres enzymes
mesurées.
![]()
La morphologie du tube digestif pourrait être modifiée après un rationnement très intense (distribution de 50 ou 60 g d'aliment par jour alors que l'ingestion est de 174g/j chez les lapins nourris librement) et bref, pratiqué pendant 1 semaine après le sevrage (de 42 à 49j d'âge; Tumova et al.): l'intestin grêle et gros intestin seraient plus longs (+10%). Néanmoins, ces résultats sont peu fiables, car les auteurs de l'étude (Chodova et al.) indiquent qu'ils ont sacrifié des animaux de poids similaires pour tous les groupes expérimentaux, c'est à dire des animaux témoins à croissance lente comparés à des lapins rationnés à croissance rapide (voir l'analyse de ce biais expérimental dans le rapport de Lebas et Duperray). Donc, comme il y a confusion des effets entre l'effet du poids vifs et celui du rationnement, il est difficile d'en tirer une conclusion sur l'effet du rationnement proprement dit.
(Krieg et al.)
|
Rapport
lignine / cellulose
|
0,26
|
0,30
|
0,32
|
0,34
|
0,40
|
Probablité
|
| - Arbocel/Vitacel (% / %) |
0
/ 8
|
2
/ 6
|
4
/ 4
|
6 / 2
|
8
/ 0
|
-
|
| - ADL (g/kg MS) |
37
|
44
|
48
|
52
|
57
|
-
|
| - Cellulose ADF-ADL (g/kg MS) |
139
|
148
|
147
|
150
|
144
|
-
|
| - Consommation (g/j sur 11 jours) |
110
|
102
|
101
|
102
|
91
|
<
0,001
|
| - GMQ (g/j sur 11 jours) |
46,7
|
46,0
|
44,5
|
44,0
|
41,2
|
=
0,042
|
| - Mobidité et mortalité / lot |
9
et 0 /40
|
8
et 1 /40
|
8
et 2 /40
|
8
et 1 /40
|
14
et 3/40
|
ns
|
| - AGV caecum (mMol /L( |
109
|
81
|
70
|
66
|
62
|
<0,001
|
| - Cryptes caecum profondeur µm |
221
|
236
|
192
|
150
|
142
|
<0,01
|
| - Cryptes caecum largeur (µm) |
36
|
53
|
40
|
39
|
14
|
<0,001
|
(Jin et Li)
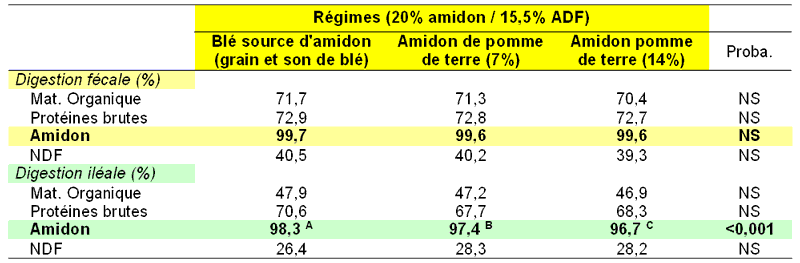
selon Trocino et al.

Ainsi, le groupe EGRAN
(Xiccato et al.) réalisé une étude inter-laboratoire
(n=5) pour préciser, avec des aliments composés et quelques
matières premières, la validité de l'analyse des
fibres totales (TDF, méthode enzymatique AOAC 991.43), et des fibres
solubles calculées par différence avec le NDF.
Les résultats indiquent une bonne répétabilité
(0,95% MS = variabilité intra-laboratoire) et une bonne reproductibilité
(1,68% % MS = variabilité inter-laboratoire) de la méthode
TDF, cette dernière étant même inférieure à
celle du NDF. Cependant, cette analyse TDF pour des ingrédients
riche en fibres "soluble" (marc de raisin ou pulpe de betterave)
montre une plus grande variabilité.
Le calcul des fibres
solubles (figure 5 ) montre un assez faible écart entre laboratoires,
dans le cas de concentration faible ou intermédiaires (Fibres solubles
entre 4 et 11% MS). Cette méthode mérite d'être harmonisée
pour améliorer sa répétabilité (2,5% MS) et
sa reproductibilité (3% MS).
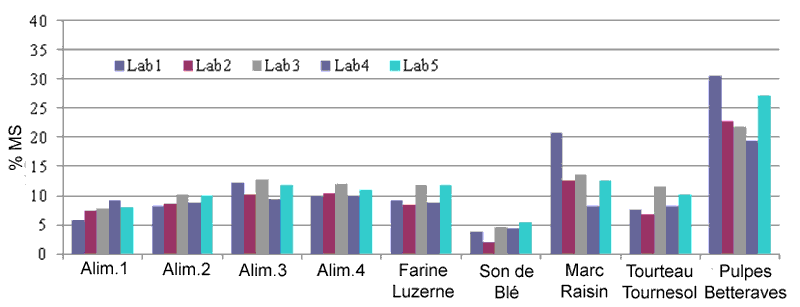
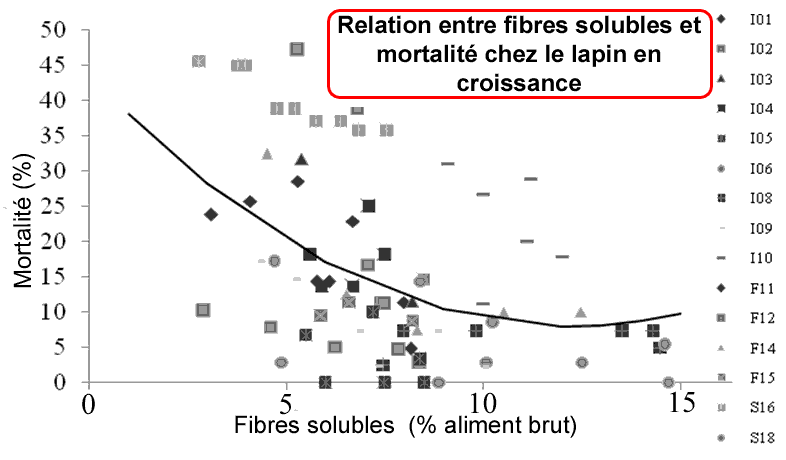
Figure 6: Relation entre fibres solubles et mortalité chez le lapin en croissance
Le rôle nutritionnel des fibres solubles "FS" chez le lapin en croissance a fait l'objet d'une intéressante synthèse (Trocino et al.), même si certains points demeurent discutables (tel que l'estimation des valeurs en "FS" d'études anciennes qui sont incluse dans leur méta-analyse bibliographique de 20 articles).
Ces auteurs proposent
des équations (basée sur 10 articles du même laboratoire)
pour corréler des paramètres de croissance, de digestion
ou de santé, avec les fibres solubles (TDF-NDF) ou avec les fibres
dites "digestibles" (FD) estimées par différence
entre TDF et ADF.
Ainsi, la digestion d'un aliment est en général un peu mieux
corrélée avec SF qu'avec FD, ce qui est logique puisque
la méta-analyse a sélectionné des études où
la concentration en fibres rapidement fermentescible varie assez fortement.
Les corrélations obtenues avec SF et FD sont semblables, pour des
critères de mortalité post-sevrage, de volumes digestifs
ou d'activité fermentaires caecales.
La relation entre la teneur en FS et l'acidité caecale ou la teneur
en AGV est globalement positive, ce qui est logique au vu de la fermentescibilité
rapide de ces composés.
L'un des points les plus intéressant est la relation avec la mortalité post-sevrage et le taux de FS (figure 6). Il semble que le taux de mortalité soit minimum entre 8 et 12% de SF (% brut). Pour des teneurs inférieures à 8 %, aucune relation n'est visible étant donné la forte variabilité de réponses observée entre études (0 à 40% de mortalité!).
Selon les auteurs, cette réduction de mortalité pourrait être associée à une meilleure "qualité" de la muqueuse (ratio villi/cryptes plus haut), et plus forte activité des disaccharidases au sevrage. Ces conclusions restent à confirmer. Les auteurs concluent également sur les recommandations en fibres pour le lapin en croissance, et tentent une comparaison un peu hasardeuse entres les recommandations de l'INRA et celles de l'université de Madrid. Néanmoins, les critères utilisés sont différents entre ces 2 "équipes". Actuellement, les recommandations de l'INRA (suites aux études du GEC) se base sur un équilibre d'apport entre l'ADF (peu digeste) et les fibres digestibles "FD" (calculées par la somme des hémicelluloses et des pectines insolubles), et n'utilise pas le critère fibres solubles calculé par différence entre TDF et NDF (trop peu fiable pour le moment).
En conclusion, quelques études ont contribué à améliorer nos connaissances sur la biocénose caecale et son activité. Il en va de même pour la synthèse sur les fibres solubles. Cependant il y a encore n écessité de poursuivre les investigations, avant de proposer de nouveaux critères de formulation basés sur les fibres rapidement fermentescibles (ou "solubles")
En revanche, les études portant sur l'impact de divers nutriments et/ou additifs sur la physiologie sont souvent assez peu originales, et quelque peu décevantes du point de vue de leur future application en cuniculture.
![]()
![]()
![]()