|

31 janvier 2017 - Journée
d'étude ASFC «Qingdao -Ombres & Lumières»
|
Génétique
au 11ème Congrès Mondial de Cuniculture
Qingdao - Chine
par
Mélanie GUNIA*
et Jacques HURTAUD**
* INRA, GenPhySE, 24 chemin de Borderouge,
31326 Castanet-Tolosan
** HYPHARM SAS, La Corbière, Roussay, 49450 Sèvremoine
|

Les 2 orateurs pendant
leur exposé |
Lors du congrès
mondial de 2016 à Qingdao en Chine, 31 communications (1 synthèse,
10 présentations orales et 20 posters) ont été
présentées dans la session génétique : 10
pour la Chine, 7 pour la France, 3 pour l'Espagne, 3 pour la Hongrie,
2 pour l'Egypte, 1 du Nigéria, 1 de Cuba, 1 de l'Equateur et
1 de Slovaquie. L'ensemble de ces contributions peut s'articuler autour
de 3 volets : génétique quantitative (14 communications),
génétique moléculaire (10 communications), caractérisation
de races (7 communications).
|
|
1
- GENETIQUE QUANTITATIVE
|
| |
1.1.
- Résistance aux maladies et longévité |

Figure 1 - Héritablité calculée et corrélations
génétiques entre caractères de santé mesurés
chez
150 000 lapins en engraissement |
Lenoir
et al. ont réalisé une étude de longévité
sur 24 000 femelles de la lignée D du sélectionneur Hycole,
en se basant sur un caractère de nombre d'IA, de la première
IA fertilisante jusqu'à la réforme ou la mort. La durée
de carrière moyenne était de 5,7 IA (80% de réforme,
20% de mort), avec un effet important de la politique de réforme
sur la fertilité et la taille de portée. L'héritabilité
du caractère de longévité était de 0,12. Les
corrélations entre ce caractère et les caractères
en sélection (nés vivants, poids de la portée à
28j, nombre de tétines, poids de la femelle adulte, et poids du
lapereau le plus léger à la naissance) étaient favorables,
ce qui a permis l'amélioration de ce caractère.
Savietto et al.
ont présenté une réflexion sur la pyramide des
âges des femelles dans les élevages. Ils se sont basés
sur les concepts de la théorie de l'histoire de vie. Les auteurs
observent que certains remplacements de femelles sont justifiés
: infertilité, animaux moins productifs, malades. Mais il y a
aussi des femelles en bonne santé qui sont remplacées
par des jeunes disponibles " de la nouvelle génétique
". Cela conduit à avoir des animaux qui investissent plus
dans les fonctions de reproduction et moins dans des fonctions d'entretien,
comme cela a été observé chez d'autres êtres
vivants. Une réduction du taux de réforme conduirait à
une vie reproductive de femelles plus longue, à une population
plus " mature " adaptée aux conditions locales, avec
en plus, dans les noyaux de sélection, une évolution vers
des lapins résistants aux maladies.
Gunia et al. ont
estimé les paramètres génétiques de signes
cliniques observés en élevage de sélection et leurs
corrélations avec des caractères de production, sur 150
000 lapins de deux lignées paternelles (Hyplus) du sélectionneur
Hypharm. Les caractères de santé étudiés
étaient les troubles digestifs non spécifiques, les troubles
respiratoires non spécifiques, et les syndromes infectieux, caractère
composite incluant les syndromes digestifs et respiratoires. Les caractères
de production étaient le poids vif et le rendement carcasse en
fin d'engraissement. Les héritabilités étaient
faibles pour les caractères de santé (de 0,01 à
0,08 ± 0,01) et modérées pour les caractères
de production. La corrélation génétique entre les
troubles digestifs et respiratoires était de -0,18 ± 0.07.
Les corrélations génétiques entre les caractères
de troubles infectieux d'une part et les troubles digestifs et respiratoires
d'autre part étaient modérées (0,65 et 0,61 ±
0,04). Les corrélations génétiques entre les caractères
de santé et de production étaient soit nulles, soit favorables.
Il est possible de sélectionner les lapins sur des symptômes
de maladies en utilisant un critère composite incluant toutes
les syndromes infectieux.
|
| |
1.2.
- Efficacité alimentaire |
| |
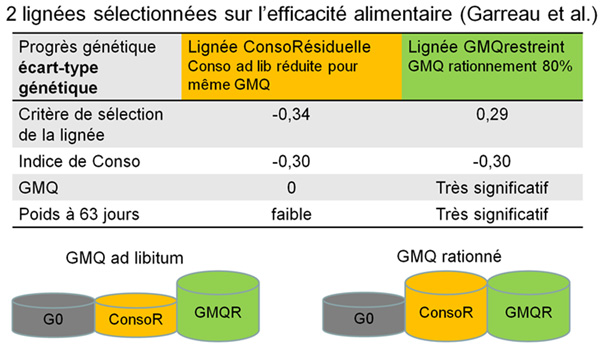
Garreau
et al. (deux communications) ont comparé 2 caractères
d'amélioration de l'efficacité alimentaire entre le sevrage
et 63 jours d'âge: 1) la consommation résiduelle (CR) afin
de sélectionner les animaux ayant une consommation ad libitum
réduite pour une même vitesse de croissance (lignée
ConsoRésiduelle), 2) le gain moyen quotidien en régime
alimentaire restreint (GMQR) afin de sélectionner les animaux
ayant une croissance élevée à ingéré
fixe (lignée GMQrestreint).
Les réponses à la sélection sont similaires pour
les deux critères de sélection (-0,34 écart-type
génétique pour la CR et 0,29 écart-type génétique
pour le GMQR). Pour l'indice de consommation (IC), la réponse
à la sélection était de -0,30 écart-type
génétique dans les deux lignées. Les réponses
à la sélection étaient nulles pour le GMQ et faible
pour le poids à 63 jours dans la lignée ConsoRésiduelle,
tandis qu'elles étaient fortement significatives dans la lignée
GMQrestreint. A la 9ème génération, les 2 lignées
sélectionnées ont été comparées à
une population témoin G0, issue d'embryons congelés, pour
les deux niveaux alimentaires, ad libitum et restreint. Quel
que soit le niveau alimentaire, les 2 lignées sélectionnées
ont un indice de consommation significativement plus faible (2,62 ±
0,02) que la lignée G0 (2,82 ± 0,02), avec toutefois des
quantités d'aliment ingéré et des poids à
63 jours différents. En régime ad libitum, la lignée
ConsoRésiduelle présente la même croissance que
la lignée G0, mais avec une moindre consommation d'aliment. En
revanche, la lignée GMQrestreint montre la plus forte croissance.
En régime restreint, les deux lignées sélectionnées
ont une croissance plus forte que la lignée G0.
|
|
|
Figure
2 - Réponses génétiques de lignées
sélectionnées pour améliorer l'efficacité
alimentaire
|
Dans
un second protocole, des lapins de la 10ème génération
de la lignée ConsoRésiduelle, ont été adoptés
par des mères de la lignée G0 à l'origine de cette
sélection, et vice versa. En parallèle, des collatéraux
étaient adoptés par des mères de leur lignée
d'origine. Les mères G10 de la lignée ConsoRésiduelle
avaient un effet défavorable sur l'IC (+0,06), quelle que soit
la lignée du jeune. Les animaux G10 pesaient moins que les G0
au sevrage (-82,9g) et à 63 jours (-161g) et avaient un GMQ (-2,36g/j)
plus faible. Ils avaient une meilleure efficacité alimentaire
avec un IC (-0,36), une CR (-548g/j) et une consommation alimentaire
totale (-839g/j) inférieurs.
Sanchez et al. ont
mené une expérimentation autour de la sélection
sur l'efficacité alimentaire en régime restreint ou ad
libitum en cages collectives. Ils ont utilisé des enregistrements
collectifs de consommation d'aliment de 6200 jeunes. L'efficacité
alimentaire était mesurée de 3 façons différentes
:
1) la consommation moyenne journalière ad libitum conditionnée
par le GMQ ad libitum (CMJAdL|GMQAdL),
2) le GMQ en régime restreint,
3) le GMQ en régime rationné conditionné par le
GMQ ad libitum (GMQR|GMQAdL).
L'héritabilité du GMQ était de 0,47 ± 0,03
en régime ad libitum et 0,40 ± 0,03 en régime
restreint, avec une variance génétique beaucoup plus faible
en régime restreint, et une corrélation génétique
de 1 entre les deux caractères. L'effet du groupe était
plus important pour les animaux en régime restreint en début
d'engraissement. Les 3 caractères d'efficacité alimentaire
avaient une héritabilité faible à modérée,
avec une variance génétique très faible pour GMQR|GMQAdL.
La corrélation génétique entre les deux caractères
CMJAdL|GMQAdL et GMQR|GMQAdL
était nulle. En conclusion, les données récoltées
en cage collective sont utilisables pour améliorer l'efficacité
alimentaire en régime ad libitum. La sélection
serait aussi potentiellement plus efficace en régime ad libitum
(à cause de la plus faible variance génétique en
régime restreint). Les différentes définitions
de l'efficacité alimentaire semblent faire appel à des
mécanismes génétiques différents.
|
| |
1.3.
Caractérisation de la carcasse : rendement et muscle |
| |
Lenoir
et al. ont étudié la possibilité d'améliorer
le rendement carcasse en utilisant de mesures in vivo effectuées
avec un échographe. Des mesures de largeur, profondeur et surface
d'un muscle (le Longissimus dorsi) ont été effectuées
à 71 jours sur 650 lapins de la lignée X du sélectionneur
Hycole. Le lendemain, le poids vif et le rendement carcasse étaient
mesurés à l'abattoir. Le caractère de surface de
muscle avait l'héritabilité la plus importante (h²=0,20)
et la corrélation génétique la plus intéressante
avec le rendement carcasse (r=0,65). Cette mesure pourrait être
utilisée en sélection pour améliorer le rendement
en carcasse.
Nagy et al. ont
présenté les paramètres génétiques
et le progrès génétique attendu pour 2 objectifs
de sélection de la lignée paternelle Pannon Large. Les
paramètres génétiques ont été estimés
sur 31 600 lapins. L'héritabilité du GMQ était
de 0,16 ± 0.01, celle du volume musculaire de la cuisse (VMC)
mesuré par tomographie était de 0,24 ± 0.03, et
celle du poids de portée à 21 jours (PP21) était
de 0,16 ± 0,04. La corrélation génétique
entre le PP21 et le GMQ (0,50 ± 0,10) était favorable,
comme celle entre le PP21 et le VMC (0,22 ± 0,12). Celle entre
GMQ et VMC était défavorable (-0,20 ± 0,08). Un
objectif de sélection avec une pondération de 23% sur
le GMQ et de 67% sur le VMC permet d'atteindre un progrès génétique
de 0,51g/jour de GMQ et 4,7 cm³ de VMC (les auteurs ne précisent
pas si ces résultats sont par génération ou par
an). Un objectif de sélection avec une pondération de
22% sur le GMQ, 50% sur le VMC et 28% sur le PP21 permet d'atteindre
un progrès génétique de 0,75g/jour de GMQ, 4,3
cm³ de VMC, et 0,06g de PP21. L'abattoir recherche avant tout un
volume de cuisse élevé, tandis que les éleveurs
veulent maintenir les performances d'élevage des femelles.
|
| |
1.4.
Caractérisation de la carcasse : taux de gras |
| |
Minguez
C. et al. ont évalué la qualité de la viande sur
258 jeunes issus de mères provenant de croisements diallèles.
Les mères provenaient de croisements entre les lignées espagnoles
A, V, H et LP. Les pères provenaient tous de la lignée R.
Le taux de gras intramusculaire, le taux de protéines et le taux
de groupes d'acides gras (monoinsaturés, polyinsaturés,
saturés) étaient mesurés dans le muscle longissimus
par NIRS (spectroscopie dans le proche infrarouge). Il n'y avait pas de
différence dans le taux de protéine entre les lignées.
La lignée A avait un taux de gras intramusculaire, des acides gras
de tous les groupes supérieurs aux autres lignées, et en
particulier supérieurs à la lignée V. La viande la
plus grasse sera donc obtenue avec la lignée A, et la viande la
plus maigre avec la lignée V. Les contrastes entre lignées
observés dépendaient principalement des effets génétiques
maternels directs (plus que des effets d'hétérosis ou d'effet
grand maternels).
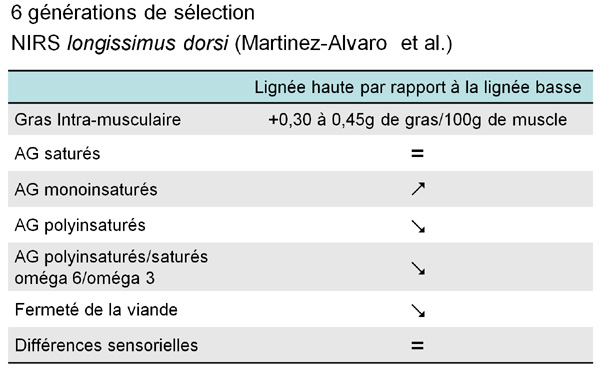
Martinez-Alvaro
et al. (deux communications) ont étudié les effets de
la sélection sur le gras intramusculaire (GI) mesuré par
NIRS (infra rouge) sur la composition en acide gras (AG). Après
6 générations de sélection divergente sur le GI,
60 lapins des 2 lignées divergentes ont été abattus
à 9 semaines d'âge, et 3 muscles ont été
récupérés (longissimus dorsi, biceps femoris,
supraspinatus). Le GI avait augmenté dans les 3 muscles,
avec une différence allant de 0,30 à 0,45g de gras/100g
de muscle entre les 2 lignées. La sélection n'a pas affecté
les AG saturés, mais a augmenté les AG monoinsaturés
et diminué les AG polyinsaturés dans la lignée
avec GI élevé. La sélection pour un GI élevé
a eu des effets défavorables sur les ratios AG polyinsaturés/
saturés et oméga6/oméga3. Martinez-Alvaro et al.
ont aussi étudié les effets de la sélection sur
le GI, sur la texture et les caractéristiques sensorielles de
la viande à partir de 116 lapins abattus à 9 et 13 semaines.
La lignée à faible GI était 10% plus ferme que
la lignée à haut GI, mais il n'y avait pas de différences
sensorielles entre les 2 lignées. La viande de lapins abattus
à 13 semaines avait un GI plus élevé qu'à
9 semaines, avec une odeur et un goût plus prononcé. Elle
était aussi plus ferme et moins juteuse.
|
|
|
Figure 3 : Sélection divergeante sur
le taux de gras intra-musculaire
|
Kasza et al. (deux
communications) ont étudié les effets d'une sélection
sur le gras corporel total sur la reproduction et la croissance. Le
taux de gras était calculé à 10 semaines en divisant
le gras corporel total mesuré par tomographie par le poids vif.
Ce taux de gras a servi de base à une sélection divergente
sur des lapins de la lignée maternelle Pannon Ka. Les auteurs
n'ont pas calculé d'héritabilité, et ne se sont
pas non plus basés sur des index de sélection dans cette
expérience. La lignée haute avait plus de nés vivants
et nés totaux que la lignée basse en 3ème génération,
avec une meilleur taille de portée à 21j, mais les deux
lignées avaient ensuite la même taille de portée
à 35j. Il n'y avait pas de différences claires entre les
lignées sur les caractères de croissance et sur la consommation
d'aliment des jeunes en engraissement lors des deux premières
générations de sélection.
|
| |
1.5.
Caractères de reproduction |
| |
Robert et al. ont
présenté les héritabilités et courbes de
progrès génétique de la lignée maternelle
Hyla C du sélectionneur Eurolap en France et en Chine. Les deux
noyaux sont connectés avec un renouvellement partiel du noyau
de sélection chinois avec des animaux français. L'héritabilité
de nombre de nés vivants était de 0,093 ± 0,009
en France et de 0,068 ± 0,013 en Chine. Les héritabilités
du poids naissance étaient similaires (0,10). Les courbes de
progrès génétique présentaient la même
pente (+0,35 nés vivants/an en France et +0,32 nés vivants
en Chine), la courbe de la Chine était en dessous de celle de
la France, ce que les auteurs attribuent à la différence
de génération. Des interactions génotype-milieu
pourraient aussi expliquer les différences entre les deux sites
de sélection.
|
|
2
- GENETIQUE MOLECULAIRE
|
| |
2.1.
Synthèse " le lapin à l'ère de la génomique
" |
| |
L. Fontanesi
a présenté cette synthèse. L'avancée des techniques
de séquençage de nouvelle génération a ouvert
l' «ère de la génomique» L'avancée majeure
pour le lapin a été le séquençage de son génome,
avec une première version en 2011, et une seconde version améliorée
(OryCun 2.0) en 2014. Lors de l'annotation du génome 19 203 gènes
codants et 3 375 gènes non-codants ont été identifiés.
Différents niveaux de variabilité entre les individus ont
été mis en évidence, avec plus de 50 millions de
marqueurs SNP (single-nucleotide polymorphism = polymorphisme d'un seul
nucléotide) et 155 marqueurs CNV (copy number variation = variabilité
du nombre de copies d'un gène). Ces données sont la base
de nombreuses études, permettant d'associer les variations du génome
avec des performances. Elles ouvrent le champ de la sélection génomique
et de l'édition de gènes.
L'approche gène
candidat consiste à choisir un gène que l'on pense être
associé à un phénotype d'intérêt (en
se basant par exemple sur des travaux dans d'autres espèces),
à identifier des polymorphismes (variations) de ce gène,
et ensuite à utiliser des méthodes statistiques permettant
de définir s'il existe une association entre les polymorphismes
du gène et l'expression du phénotype. L'approche gène
candidat est rapide mais a ses faiblesses, dues au choix à priori
du gène à étudier. Par exemple, l'effet de mutations
du gène de la myostatine, très important dans de nombreuses
espèces sur les caractères de carcasse, est très
faible voire inexistant en lapin. L. Fontanesi liste les gènes
candidats étudiés en lapin associés avec la croissance
et la production de viande, les caractères de reproduction, les
caractères de résistance aux maladies et les caractères
liés au pelage.
Les analyses de
détection de QTL, très nombreuses dans d'autres populations
animales, ont jusqu'à présent été rares
en lapin, avec une seule étude publiée avec des marqueurs
microsatellites portant sur des caractères de carcasse. Avec
la découverte de millions de marqueurs SNP et la disponibilité
des nouvelles technologies, cette situation est vouée à
changer.
Toutes les conditions
sont réunies pour le développement de la sélection
génomique en lapin, avec le séquençage du lapin,
le développement d'un panel commercial de SNP pour le génotypage,
la disponibilité de méthodes statistiques développées
pour d'autres espèces et de stratégies (notamment en porc)
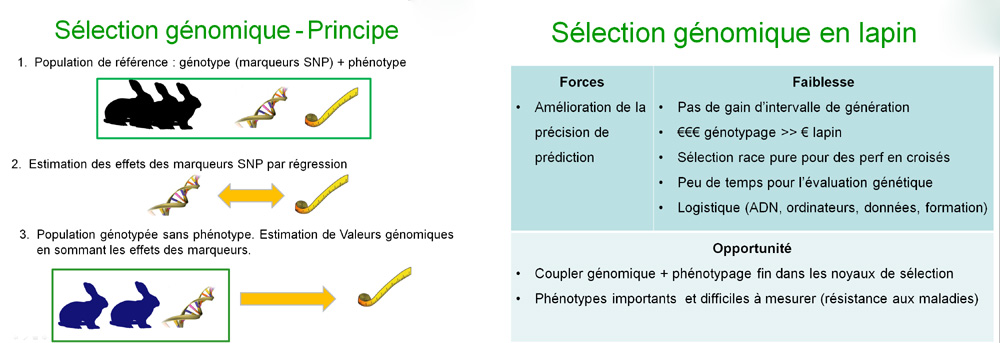
qui pourraient être adaptées au lapin. La sélection
génomique est basée sur la prédiction de Valeurs
Génomiques (GEBV) obtenues pour chaque individu en sommant tous
les effets des SNP sur l'ensemble de son génome. Les effets des
marqueurs sont estimés par régression des phénotypes
sur les génotypes sur une population de référence
(pour qui on dispose à la fois de l'information des phénotypes
et des génotypes). L'estimation de ces effets permet ensuite
de prédire les Valeurs Génomiques d'animaux avec génotypes
mais sans phénotypes.
La sélection
génomique permet en principe d'accroitre le progrès génétique
en réduisant l'intervalle de génération et en améliorant
la précision de prédiction. En lapin comme en porc, il
semblerait que les avantages ne viennent que de l'amélioration
de la précision. Cet avantage est contrebalancé par les
limites propres au système d'élevage de lapin : 1) coût
élevé du génotypage comparé à la
valeur d'un individu, 2) schémas pyramidaux avec une sélection
en race pure pour des performances finales exprimés par des animaux
croisés, 3) peu de temps disponible pour l'évaluation
génétique, 4) la mise en place des aspects logistiques
(stockage d'ADN, puissance de calcul des machines, manipulation des
données, formation des personnes). Cependant, il pourrait être
possible de combiner des approches de sélection génomique
et de phénotypage fin dans les noyaux de sélection pour
des phénotypes importants et difficiles à mesurer (comme
la résistance aux maladies).
|
| |
|
| |
2.2.
Etudes de gènes |
| |
Wang
L et al. ont utilisé une approche gène candidat pour étudier
l'association de polymorphisme des gènes PIK3CA et AKT3 sur la
croissance des lapins. Ils ont séquencés ces gènes
pour identifier leurs mutations et ont comparé les fréquences
de ces mutations dans différentes populations. Certaines mutations
ont un effet sur la croissance (qui pouvait être meilleure, mais
aussi conduire à une croissance pathologique de certains tissus).
Les effets de ces gènes sur les protéines et leurs liens
avec d'autres gènes n'ont pas été étudiés.
Wang J et al. ont
également utilisé une approche gène candidat pour
étudier les polymorphismes du gène de la calpastatine
dans 3 race de lapins. Certaines variations de ces gènes semblent
être associées avec un taux de gras intramusculaire plus
élevé.
Ondruska et al. ont
étudié le polymorphisme du promoteur du gène de
la protéine-C-réactive, protéine qui a un rôle
dans l'immunité innée, sur des lignées à
haute et basse variabilité du nombre de nés vivants. Il
y aurait une relation entre hétérozygotie de ce gène
et viabilité des jeunes au servage.
Chen et al. ont
séquencé le gène codant pour la protéine
NRAMP1 chez le lapin Fujian Jaune. Cette protéine est connue
pour avoir un rôle dans la sensibilité aux maladies infectieuses
dans d'autres espèces. Ce gène pourrait servir de gène
candidat pour l'étude de la résistance aux maladies chez
le lapin.
Zhang et al. ont
étudié le polymorphisme de plusieurs gènes codants
pour les récepteurs de type Toll (TLR) intervenant dans l'immunité
innée.
Sang et al. ont
séquencé le gène codant pour l'inhibine beta A
chez le lapin de Fujian jaune.
Fu et al. ont étudié
l'expression de deux gènes, GPR41 et GPR43, connus pour avoir
un effet sur les acides gras à courte chaine.
Kuang et al. ont
étudié l'expression de deux gènes, FAS et COT1,
codant pour des enzymes ayant un rôle dans le dépôt
adipeux.
|
| |
2.3.
Profil du microbiome intestinal |
| |
Deng et al. ont
décrit des profils de microbiome intestinal en utilisant un séquençage
16s.
|
|
3.
CARACTERISATION DE RACES
|
| |
3.1.
Performances zootechniques de différentes races |
| |
Chen et al. et Xie
et al. ont décrit les performances des races chinoises Minxinnan
Noir et Fujian Blanc. Oseni et al. ont décrit les performances
de lapins Néo-Zelandais blancs au Nigéria.
|
| |
3.2.
Caractérisation de populations au niveau moléculaire |
| |
Emam
et al. (deux communications) ont étudié la phylogénie
de 3 races locales égyptiennes (Baladi Rouge, Baladi Noir et Gabali
du Sinai) et des lapins espagnols (domestiques et sauvages) en se basant
sur des marqueurs microsatellites et le séquençage d'un
gène mitochondrial.
Orheruata et al.
ont étudié au Nigéria la distance génétique
entre des population de Neo Zelandais, Californien, Rex, Hollandais
et une population composite en se basant sur une technique RAPD (random
amplified polymorphic DNA = amplification aléatoire d'ADN polymorphe).
Ils ont montré qu'il existait peu de diversité dans les
populations standards, et beaucoup de variabilité dans la population
composite qui peut être un bon support pour la sélection.
|
| |
3.3.
Caractérisation d'hybrides issus de 4 races |
| |
Ponce de Leon et
al. ont étudié à Cuba 6 300 lapereaux en pré-sevrage,
issus des croisements diallèles entre 4 races : le Californien
(C), le Chinchilla (Ch), le Néo-Zelandais Blanc (Nz) et le Semi-Géant
Blanc (S). Tous les parents ont été croisés pour
produire des hybrides de toutes les combinaisons. Les caractères
observés étaient les nés totaux, nés vivants,
viabilité à la naissance, nombre de sevrés, proportion
de portées sevrées, viabilité au sevrage. Les femelles
S et Nz étaient les meilleures en viabilité et nombre
de sevrés. Les meilleurs croisements étaient SNz, ChS,
ChNz, CS, avec S et Nz comme lignées maternelles.
|
|
4
- CONCLUSION
|
| |
Les
caractères de longévité et résistance aux
maladies sont bien représentés, avec en arrière-plan
les problématiques liées à la réduction des
intrants médicamenteux en élevage.
Les caractères d'efficacité alimentaire et de qualité
de carcasse ont occupé une place importante dans les communications
de génétique. Il existe en effet de nombreuses façons
de mesurer et améliorer l'efficacité alimentaire, avec des
résultats variés. De même, il existe de nombreux axes
d'amélioration de la carcasse : amélioration du rendement
carcasse, du volume de cuisse, du gras intramusculaire, voire du gras
total. Les techniques de mesures in vivo (échographie, tomographie,
spectroscopie dans le proche infrarouge) sont devenues beaucoup plus accessibles
au monde de l'élevage et fournissent de nouveaux phénotypes,
très fins, qui ouvrent de nouvelles portes à la sélection.
Et, comme annoncé dans la synthèse, l'ère de la génomique
s'ouvre pour le lapin, avec le séquençage de son génome
et la construction d'une puce SNP. La combinaison des nouveaux outils
de génotypage et de phénotypage va entrainer des changements
majeurs dans les méthodes de sélection du lapin. |
| |
|
|
|